Онто и филогенез скелетной мускулатуры животных. Лекция миология филогенез онтогенез и функциональная анатомия мышечной. Мышца: определение, строение
Не изолированная мышечная система
Единый кожно-мускульный мешок
Появление поперечно-полосатой мышечной ткани
Разделение мышечных тяжей на миотомы
Развитие групп мышц
Развитие мышц конечностей (изменение среды обитания)
Развитие диафрагмы
Развитие всех групп мышц - выполнение дифференцированных движений
2 Онтогенез мышечной системы: источники и сроки развития.
Производные миотома: из дорзального участка развиваются мышцы спины
из вентрального - мышцы груди и живота
Мезенхима - мышцы конечностей
I висцеральная дуга (ВД) - жевательные мышцы
II ВД - мимические мышцы
III и IV ВД - мышцы мягкого неба, глотки, гортани, верхней части пищевода
V ВД - грудино-ключично-сосцевидная и трапециевидная мышцы
Из затылочных миотомов - мышцы языка
Из предушных миотомов - мышцы глазного яблока
Мышцы развиваются из мезодермы . На туловище они возникают из первичной сегментированной мезодермы – сомитов : 3-5 затылочных, 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых, 4-5 копчиковых.
Каждый сомит подразделяется на склеротом, дерматом и миотом – из него и развиваются мышцы туловища. Сомиты появляются рано, когда длина зародыша составляет 10-15 мм.
Из дорсальных частей миотомов возникают глубокие, собственные (аутохтонные) мышцы спины, из вентральных – глубокие мышцы груди и живота. Они закладываются, развиваются и остаются в пределах туловища – поэтому называются аутохтонными (местными, туземными ). Очень рано миотомы связываются с нервной системой и каждому мышечному сегменту соответствует нервный сегмент. Каждый нерв следует за развивающейся мышцей, врастает в нее и, пока она не дифференцировалась, подчиняет своему влиянию.
В процессе развития часть скелетных мышц перемещается с туловища и шеи на конечности – трункофугальные мышцы: трапециевидная, грудино-ключично-сосцевидная, ромбовидные, поднимающая лопатку и др. Часть мышц, наоборот, направляется с конечностей на туловище – трункопетальные мышцы: широчайшая мышца спины, большая и малая грудные, большая поясничная.
Мышцы головы мимические и жевательные, над- и подъязычные мышцы шеи развиваются из несегментированной вентральной мезодермы, входящей в состав висцеральных (жаберных) дуг. Они называются висцеральными и, например, жевательные мышцы развиваются на основе первой висцеральной дуги, а мимические – второй. Однако мышцы глазного яблока и языка развиваются из затылочных миотомов сегментированной мезодермы. Глубокие передние и задние мышцы шеи также возникают из затылочных шейных миотомов, а поверхностная и средняя группа мышц в передней области шеи развиваются на основе несегментированной мезодермы висцеральных дуг.
3 Мышца: определение, строение.
Мускул (мышца) – орган, построенный из мышечных волокон (клеток), каждое из них обладает соединительно-тканной оболочкой – эндомизием . В пучки мышечные волокна объединяет другая фиброзная оболочка – перимизий , а весь мускул заключается в общую фиброзную оболочку, образованную фасцией – эпимизий . Между пучками проходят сосуды и нервы, снабжающие мышечные волокна.
На макроуровне скелетная мышца имеет:
брюшко (вентер) – мясистая часть органа, занимающая его средину;
сухожилие (тендо), относящееся к дистальному концу, оно может быть в виде апоневроза, сухожильных перемычек, длинных пучков продольных фиброзных волокон;
головку , составляющую проксимальную часть;
сухожилие и головка крепятся на противоположных концах костей.
Проксимальное сухожилие или головка мышцы - начало мышцы на кости находится ближе к срединной оси тела – это фиксированная точка (punctum fixum) (как правило совпадает с началом мышцы). Дистальное сухожилие, «хвост» - конец мышцы лежит на кости дистальнее и, являясь местом прикрепления, называется подвижной точкой (punctom mobile). При сокращении мышцы точки сближаются, а при изменении положения тела могут меняться местами.
Сухожилия разные по форме: тонкие длинные сухожилия имеют мышцы конечностей; мышцы, участвующие в формировании стенок брюшной полости, имеют широкое плоское сухожилие, расположенное меж двумя брюшками - сухожильное растяжение или апоневроз.
Филогенез систем органов хордовых рассмотрен в соответствии с прогрессивным направлением эволюции этого типа животных от подтипа Бесчерепные до класса Млекопитающие. Организация систем органов класса Птицы не описана в связи с тем, что птицы произошли от пресмыкающихся значительно позже млекопитающих и являются боковой ветвью эволюции хордовых.
Наружные покровы
Покровы любых животных всегда выполняют функцию восприятия внешних раздражении, а также защищают тело от вредных воздействий среды. Интенсификация первой функции покровов приводит в процессе эволюции многоклеточных животных к возникновению нервной системы и органов чувств. Интенсификация второй функции сопровождается дифференцировкой. Характерно также расширение функций, в результате чего кожа как орган защиты участвует также в газообмене, терморегуляции и выделении, выкармливании потомства. Это связано с усложнением строения слоев кожи, появлением и дальнейшим преобразованием многочисленных придатков и желез.
У всех хордовых кожа имеет двойное - экто- и мезодермальное - происхождение. Из эктодермы развивается эпидермис, из мезодермы - дерма. Для бесчерепных характерна слабая степень дифференцированности обоих слоев кожи. Эпидермис однослойный цилиндрический, содержащий одноклеточные слизистые железы, дерма рыхлая, содержит небольшое количество соединительнотканных клеток.
В подтипе Позвоночные эпидермис становится многослойным, причем в нижнем слое клетки постоянно размножаются, а в верхних слоях - дифференцируются, гибнут и слущиваются. В дерме появляются соединительнотканные волокна, придающие покровам прочность. Кожа образует придатки, многообразные в зависимости от образа жизни и уровня организации, а также железы, выполняющие различные функции.
У рыб в эпидермисе железы одноклеточные. Как и у ланцетника, они выделяют слизь, облегчающую движения в воде. Тело рыб покрыто чешуей, имеющей разное строение в зависимости от их систематического положения. Чешую хрящевых рыб называют плакоидной. Она имеет форму шипа и состоит из дентина, покрытого снаружи эмалью (рис. 14.1). Дентин мезодермального происхождения, он образуется за счет функционирования соединительнотканных клеток, выпячивающихся снаружи в виде сосочка. Эмаль, представляющая собой более твердое, чем дентин, неклеточное вещество, формируется сосочком эпидермиса и покрывает плакоидную чешую снаружи.
Вся поверхность тела хрящевых рыб, а также ротовая полость, слизистая оболочка которой происходит из эктодермы, покрыты плакоидной чешуей. Естественно, что функции чешуи в ротовой полости связаны с захватом и удержанием пищи, поэтому они сильно увеличены и являются зубами. У костных рыб чешуя иного типа. Она имеет вид тонких круглых костных пластинок, покрытых тонким слоем эпидермиса. Костная чешуя развивается полностью за счет дермы, но по происхождению связана с примитивной плакоидной.
Кожа примитивных вымерших земноводных - стегоцефалов - соответствовала покровам рыб и тоже была покрыта чешуей. Современные амфибии имеют тонкую гладкую кожу без чешуи, принимающую участие в газообмене. Этому способствует наличие большого количества многоклеточных слизистых желез, секрет которых постоянно увлажняет покровы и обладает бактерицидными свойствами. Некоторые кожные железы ряда земноводных дифференцировались в органы-продуценты токсинов, защищающих их от врагов (см. разд. 23.1).
Рис. 14.1. Закладка плакоидной чешуи:
1 -клетки-эмалеобразователи, 2- эпидермис, 3- эмаль, 4- склеробласты-дентинообразователи, 5- дентин, 6- сосочек дермы
Пресмыкающиеся, перешедшие полностью к наземному существованию, имеют сухую кожу, не участвующую в дыхании. Верхний слой эпидермиса ороговевает. Роговые чешуйки у некоторых рептилий тонки и эластичны, у других-сливаются вместе, образуя, как у черепах, мощный роговой панцирь. Большинство пресмыкающихся по мере роста линяют, сбрасывая периодически свой роговой покров. Современные пресмыкающиеся не имеют кожных желез.
Кожные покровы млекопитающих построены наиболее сложно в связи с выполнением ими многообразных функций. Характерны различные производные кожи: волосы, когти, рога, копыта, а также потовые, сальные и млечные железы. Более примитивные млекопитающие - насекомоядные, грызуны и некоторые другие - наряду с волосяным покровом сохранили также роговые чешуи на хвосте. Волосы у них растут в промежутках между чешуйками, группами по 3-7. У более прогрессивных млекопитающих, утративших чешуи, сохраняется такое же расположение волос (рис. 14.2), покрывающих практически все тело, кроме некоторых участков, например подошв и ладоней у человека.
Волосы многих млекопитающих дифференцированы на типичные, служащие для терморегуляции, и большие, или вибриссы , основания которых связаны с чувствительными нервными окончаниями. У большинства млекопитающих вибриссы расположены в области рта и носа, у приматов они редуцированы в связи с усилением осязательной функции передних конечностей, у многих яйцекладущих и сумчатых - разбросаны по всему телу. Этот факт может свидетельствовать о том, что волосяной покров предков млекопитающих первично выполнял осязательные функции, а затем, по мере увеличения количества волос, стал принимать участие в терморегуляции. В онтогенезе человека закладывается большее количество волосяных зачатков, но к концу эмбриогенеза наступает редукция большинства из них.
Потовые железы млекопитающих гомологичны кожным железам амфибий. Их секрет может быть слизистым, содержать белки и жир. Некоторые потовые железы дифференцировались у ранних млекопитающих в млечные железы. У яйцекладущих (утконос, ехидна) млечные железы сходны с потовыми по строению и развитию. По краям развивающегося соска молочной железы можно обнаружить последовательные переходы от типичных потовых к млечным железам (рис. 14.3). Количество млечных желез и сосков коррелирует с плодовитостью (от 25 до одной пары), но в эмбриогенезе всех млекопитающих на брюшной поверхности закладываются «млечные линии», тянущиеся от подмышечной впадины до паха. Впоследствии на этих линиях дифференцируются соски, большая часть которых затем подвергается редукции и исчезает. Так, в эмбриогенезе человека закладывается вначале пять пар сосков, а впоследствии остается лишь одна (рис. 14.4).

Рис. 14.4. Эмбриогенез передней брюшной стенки человека. А - зародыш в возрасте 5 недель (видны млечные линии); Б - дифференцировка пяти пар сосков; В - зародыш в возрасте 7 недель

Рис. 14.5. Атавистические аномалии развития кожи.
А - гипертрихоз; Б - полимастия
Сальные железы образуются в коже только у млекопитающих. Их секрет, смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность.
Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуи, характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития: гипертрихоз (повышенное оволосение), полителию (увеличенное количество сосков), полимастию (увеличенное количество млечных желез) (рис. 14.5). Все они связаны с нарушением редукции избыточного количества этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами - млекопитающими. Именно поэтому у человека и других млекопитающих невозможно рождение потомства с атавистическими признаками кожных покровов, характерными для более отдаленных предков. Одним из самых известных признаков недоношенности новорожденных является повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Филогенез двигательной функции лежит в основе прогрессивной эволюции животных. Поэтому уровень их организации в первую очередь зависит от характера двигательной активности, которая определяется особенностями организации опорно-двигательного аппарата, претерпевшего в типе Хордовые большие эволюционные преобразования в связи со сменой сред обитания и изменения форм локомоции. Действительно, водная среда у животных, не имеющих наружного скелета, предполагает однообразные движения за счет изгибов всего тела, в то время как жизнь на суше более способствует их перемещению с помощью конечностей.
Рассмотрим в отдельности эволюцию скелета и мышечной системы.
Скелет
У хордовых скелет внутренний. По строению и функциям подразделяется на осевой, скелет конечностей и головы.
Осевой скелет
В подтипе Бесчерепные имеется только осевой скелет в виде хорды. Она построена из сильно вакуолизированных клеток, плотно прилегающих друг к другу и покрытых снаружи общими эластической и волокнистой оболочками. Упругость хорде придают тургорное давление ее клеток и прочность оболочек. Хорда закладывается в онтогенезе всех хордовых и выполняет у более высокоорганизованных животных не столько опорную, сколько морфогенетическую функцию, являясь органом, осуществляющим эмбриональную индукцию.
На протяжении всей жизни у позвоночных хорда сохраняется только у круглоротых и некоторых низших рыб. У всех остальных животных она редуцируется. У человека в постэмбриональном периоде сохраняются рудименты хорды в виде nucleus pulposus межпозвоночных дисков. Сохранение избыточного количества хордального материала при нарушении его редукции чревато возможностью развития у человека опухолей - хордом, возникающих на его основе.
У всех позвоночных хорда постепенно вытесняется позвонками, развивающимися из склеротомов сомитов, и функционально заменяется позвоночным столбом. Это один из выраженных примеров гомотопной субституции органов (см. § 13.4). Формирование позвонков в филогенезе начинается с развития их дуг, охватывающих нервную трубку и становящихся местами прикрепления мышц. Начиная с хрящевых рыб обнаруживается охрящевение оболочки хорды и разрастание оснований позвонковых дуг, в результате чего формируются тела позвонков. Срастание верхних позвонковых дуг над нервной трубкой образует остистые отростки и позвоночный канал, в который заключена нервная трубка (рис. 14.6).

Рис. 14.6. Развитие позвонка. А-ранний этап; Б- последующая стадия:
1 -хорда, 2- оболочка хорды, 3- верхние и нижние позвонковые дуги, 4- остистый отросток, 5- зоны окостенения, 6-рудимент хорды, 7- хрящевое тело позвонка
Замещение хорды позвоночным столбом - более мощным органом опоры, имеющим сегментарное строение,- позволяет увеличить общие размеры тела и активизирует двигательную функцию. Дальнейшие прогрессивные изменения позвоночного столба связаны с тканевой субституцией - заменой хрящевой ткани на костную, что обнаруживается у костных рыб, а также с дифференцировкой его на отделы.
У рыб только два отдела позвоночника: туловищный и хвостовой. Это связано с перемещением их в воде за счет изгибов тела.
Земноводные приобретают также шейный и крестцовый отделы, представленные каждый одним позвонком. Первый обеспечивает большую подвижность головы, а второй - опору задним конечностям.
У пресмыкающихся удлиняется шейный отдел позвоночника, первые два позвонка которого подвижно соединены с черепом и обеспечивают большую подвижность головы. Появляется поясничный отдел, еще слабо отграниченный от грудного, а крестец состоит уже из двух позвонков.
Млекопитающие характеризуются стабильным количеством позвонков в шейном отделе, равным 7. В связи с большим значением в движении задних конечностей крестец образован 5-10 позвонками. Поясничный и грудной отделы четко отграничены друг от друга.
У рыб все туловищные позвонки несут ребра, не срастающиеся друг с другом и с грудиной. Они придают телу устойчивую форму и обеспечивают опору мышцам, изгибающим тело в горизонтальной плоскости. Эта функция ребер сохраняется у всех позвоночных, совершающих змеевидные движения,- у хвостатых земноводных и пресмыкающихся, поэтому у них ребра также располагаются на всех позвонках, кроме хвостовых.
У пресмыкающихся часть ребер грудного отдела срастается с грудиной, формируя грудную клетку, а у млекопитающих в состав грудной клетки входит 12-13 пар ребер.

Рис. 14.7. Аномалии развития осевого скелета. А - рудиментарные шейные ребра (показаны стрелками); Б - несращение остистых отростков позвонков в грудной и поясничной областях. Спинномозговые грыжи
Онтогенез осевого скелета человека рекапитулирует основные филогенетические стадии его становления: в периоде нейруляции закладывается хорда, заменяющаяся впоследствии хрящевым, а затем и костным позвоночником. На шейных, грудных и поясничных позвонках развивается по паре ребер, после чего шейные и поясничные ребра редуцируются, а грудные срастаются спереди друг с другом и с грудиной, формируя грудную клетку.
Нарушение онтогенеза осевого скелета у человека может выразиться в таких атавистических пороках развития, как несрастание остистых отростков позвонков, в результате чего формируется spinabifida - дефект позвоночного канала. При этом часто через дефект выпячиваются мозговые оболочки и образуется спинномозговая грыжа (рис. 14.7).
В возрасте 1,5-3 мес. зародыш человека обладает хвостовым отделом позвоночника, состоящим из 8-11 позвонков. Нарушение их редукции в последующем объясняет возможность возникновения такой известной аномалии осевого скелета, как персистирование хвоста.
Нарушение редукции шейных и поясничных ребер лежит в основе их сохранения в постнатальном онтогенезе.
Скелет головы
Продолжением осевого скелета спереди является осевой, или мозговой, череп, служащий для защиты головного мозга и органов чувств. Рядом с ним развивается висцеральный, или лицевой череп, образующий опору передней части пищеварительной трубки. Обе части черепа развиваются по-разному и из разных зачатков. На ранних этапах эволюции и онтогенеза они не связаны между собой, но позже эта связь возникает.

Рис. 14.8. Череп человека с методическим швом (указан стрелкой)
В задней части осевого черепа в процессе развития обнаруживаются следы сегментации, поэтому считают, что он представляет собой результат слияния друг с другом закладок передних позвонков. В состав мозгового черепа включаются также закладки хрящевых капсул мезенхимального происхождения, окружающие органы слуха, обоняния и зрения. Кроме того, часть мозгового черепа (лежащая кпереди от турецкого седла), не имеющая сегментации, развивается, по-видимому, как новообразование в связи с увеличением размеров переднего мозга.
Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую и костную.
У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части.
Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю.
У костных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания (основная, клиновидная, решетчатая кости) и за счет возникновения покровных костей в верхней его части (теменные, лобные, носовые кости). Кости осевого черепа в процессе прогрессивной эволюции претерпевают олигомеризацию. Появление большого количества зон окостенения и последующее слияние их вместе при формировании таких костей, как лобная, височная и др., свидетельствуют об этом. Широко известны у человека такие аномалии мозгового черепа, как наличие межтеменных, а также двух лобных костей с метопическим швом между ними (рис. 14.8). Никакими патологическими явлениями они не сопровождаются и обнаруживаются поэтому обычно случайно после смерти.
Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной дугам взрослых животных. Следующие дуги в числе 4-5 пар выполняют опорную функцию для жабр и называются жаберными.
У хрящевых рыб впереди челюстной дуги располагаются обычно еще 1-2 пары предчелюстных дуг, имеющих рудиментарный характер. Это свидетельствует о том, что у предков позвоночных имелось большее количество висцеральных дуг, чем б или 7, а их дифференцировка происходила на фоне олигомеризации.
Челюстная дуга состоит из двух хрящей. Верхний называют нёбно-квадратным, он выполняет функцию первичной верхней челюсти. Нижний, или меккелев, хрящ - первичная нижняя челюсть. На вентральной стороне глотки меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибулярного хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвеска к мозговому черепу. Такой тип соединения челюстей и осевого черепа называют гиостильным (рис. 14.9).
У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей - челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп остается гиостильным.
Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом - нёбно-квадратным хрящом - срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным. Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвеска, располагаясь в области первой жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки - столбика,- передающей звуковые колебания от наружного к внутреннему уху.
Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи.
Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения.
Одна слуховая косточка - столбик,- характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих (рис. 14.9).

Рис. 14.9. Эволюция двух первых висцеральных жаберных дуг позвоночных.
А- хрящевая рыба; Б- земноводное; В- пресмыкающееся; Г- млекопитающее:
1 -нёбно-квадратный хрящ, 2-меккелев хрящ, 3- гиомандибулярный хрящ, 4-гиоид, 5- столбик, 6- накладные кости вторичных челюстей, 7-наковаленка, 8- стремечко, 9- молоточек; гомологичные образования обозначены соответствующей штриховкой
Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки является механизмом формирования такого порока развития среднего уха, как расположение в барабанной полости только одной слуховой косточки - столбика, что соответствует строению звукопередающего аппарата земноводных и пресмыкающихся.
Скелет конечностей
У хордовых выделяются непарные и парные конечности. Непарные (спинные, хвостовой" и анальный плавники) являются основными органами передвижения у бесчерепных, рыб и в меньшей степени у хвостатых амфибий. У рыб возникают также парные конечности - грудные и брюшные плавники, на базе которых впоследствии развиваются парные конечности наземных четвероногих животных.
Подробнее рассмотрим происхождение и эволюцию парных конечностей.
У личинок рыб, а также у современных бесчерепных вдоль тела с обеих сторон тянутся боковые кожные складки, называемые метаплевральными (рис. 14.10). Они не имеют ни скелета, ни собственной мускулатуры, выполняя пассивную роль - стабилизацию положения тела и увеличение площади брюшной поверхности, облегчающие перемещение в водной среде. Вероятно, у предков рыб, переходящих к более активному образу жизни, в этих складках появились мышечные элементы и хрящевые лучи, связанные с сомитами по происхождению и поэтому расположенные метамерно. Такие складки, приобретя подвижность, могут выполнять роль рулей глубины, однако для изменения положения тела в пространстве большее значение имеют их передние и задние отделы, как наиболее удаленные от центра тяжести. Поэтому эволюция шла по пути интенсификации функций крайних отделов и ослабления функций центральных частей.

Рис. 14.10. Формирование передних и задних конечностей из метаплевральных складок: I -III -гипотетические этапы эволюции
В результате из передних отделов складок развились грудные, а из задних - брюшные плавники (рис. 14.10). Не исключено, что формированию только двух пар конечностей на боковых сторонах тела предшествовал распад сплошных складок на ряд парных плавников, большее значение из которых также имели передние и задние. Об этом свидетельствует существование ископаемых остатков древнейших низкоорганизованных рыб с многочисленными плавниками (рис. 14.11). За счет слияния оснований хрящевых лучей возникли плечевой и тазовый пояса. Остальные их участки дифференцировались в скелет свободных конечностей.

Рис. 14.11. Древняя акулообразная рыба с многочисленными парными конечностями
У большинства рыб в скелете парных плавников выделяют проксимальный отдел, состоящий из небольшого числа хрящевых или костных пластинок, и дистальный, в состав которого входит большое количество расположенных радиально сегментированных лучей. С поясами конечностей плавники соединены малоподвижно. Они не могут служить опорой телу при передвижении по дну или суше. У кистеперых рыб скелет парных конечностей имеет иное строение. Общее количество их костных элементов уменьшено, и они имеют более крупные размеры. Проксимальный отдел состоит только из одного крупного костного элемента, соответствующего плечевой или бедренной костям передних или задних конечностей. Далее следуют две более мелкие косточки, гомологичные локтевой и лучевой или большой и малой берцовым костям. На них опираются 7-12 радиально расположенных лучей. В соединении с поясами конечностей у такого плавника участвуют только гомологи плечевой или бедренной костей, поэтому плавники кистеперых рыб оказываются активно подвижными (рис. 14.12, А, Б ) и могут использоваться не только для изменения направления движения в воде, но и для перемещения по твердому субстрату.
Жизнь этих рыб в мелких пересыхающих водоемах в девонском периоде способствовала отбору форм с более развитыми и подвижными конечностями. Наличие у них добавочных органов дыхания (см. разд. 14.3.4) стало второй предпосылкой выхода на сушу и возникновения других адаптации к наземному существованию, результатом чего явилось происхождение земноводных и всей группы Tetrapoda. Первые их представители - стегоцефалы - обладали семи- и пятипалыми конечностями, сохраняющими сходство с плавниками кистеперых рыб (рис. 14.12, B )

Рис. 14.12. Скелет конечности кистеперой рыбы (А ), его основание (Б ) и скелет передней лапы стегоцефала (В ): I- плечевая кость, 2-локтевая кость, 3- лучевая кость
В скелете запястья сохранено правильное радиальное расположение костных элементов в 3-4 ряда, в пястье располагается 7-5 костей, а далее также радиально лежат фаланги 7-5 пальцев.
У современных земноводных количество пальцев в конечностях равно пяти или происходит их олигомеризация до четырех.
Дальнейшее прогрессивное преобразование конечностей выражается в увеличении степени подвижности соединений костей, в уменьшении количества костей в запястье сначала до трех рядов у амфибий и затем до двух - у пресмыкающихся и млекопитающих. Параллельно уменьшается также и количество фаланг пальцев. Характерно также удлинение проксимальных отделов конечности и укорочение дистальных.
Расположение конечностей в ходе эволюции также меняется. Если у рыб грудные плавники находятся на уровне первого позвонка и обращены в стороны, то у наземных позвоночных в результате усложнения ориентации в пространстве появляется шея и возникает подвижность головы, а у пресмыкающихся и особенно у млекопитающих в связи с приподнятием тела над землей передние конечности перемещаются кзади и ориентируются не горизонтально, а вертикально. То же касается и задних конечностей.
Многообразие условий обитания, предоставляемых наземным образом жизни, обеспечивает многообразие форм передвижения: прыжки, бег, ползание, полет, рытье, лазание по скалам и деревьям, а при возвращении в водную среду - и плавание. Поэтому у наземных позвоночных можно встретить как почти неограниченное многообразие конечностей, так и их полную вторичную редукцию, причем многие сходные адаптации конечностей в разнообразных средах многократно возникали конвергентно (рис. 14.13). Однако в процессе онтогенеза у большинства наземных позвоночных проявляются общие черты в развитии конечностей: закладка их зачатков в виде малодифференцированных складок, формирование в кисти и стопе вначале шести или семи зачатков пальцев, крайние из которых вскоре редуцируются и в дальнейшем развивается только пять (рис. 14.14).

Рис. 14.13. Скелет передней конечности наземных позвоночных. А -лягушка- Б -саламандра; В -крокодил; Г -летучая мышь; Д -человек: 1 -плечевая кость, 2-лучевая кость, 3 -кости запястья, 4 -пясти, 5 -фаланги пальцев, 6 -локтевая кость
Рис. 14.14. Строение развивающейся конечности позвоночного: рр - prepollex, pin - postminimus - дополнительные рудиментарные I и VII пальцы
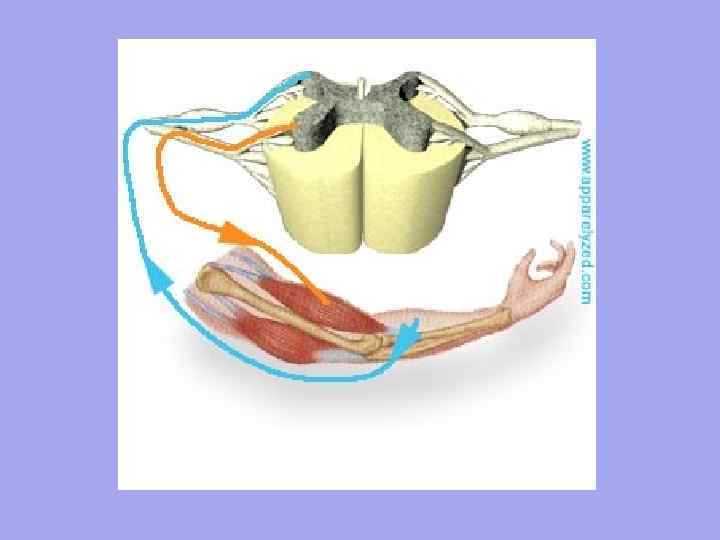
Интересно, что в эмбриогенезе высших позвоночных рекапитулирует не только строение конечностей предков, но и процесс их гетеротопии. Так, у человека верхние конечности закладываются на уровне 3-4-го шейных позвонков, а нижние - на уровне поясничных позвонков. В это же время конечности получают иннервацию из соответствующих отделов спинного мозга. Гетеротопия конечностей сопровождается формированием шейного, поясничного и крестцового нервных сплетений, нервы которых связаны с одной стороны с теми сегментами спинного мозга, из которых они вырастали в момент формирования конечностей, а с другой - с конечностями, переместившимися на новое место (рис. 14.15; см. также разд. 14.2.2.2).
В онтогенезе человека возможны многочисленные нарушения, ведущие к формированию врожденных пороков развития конечностей атавистического плана. Так, полидактилия, или увеличение количества пальцев, наследующаяся как аутосомно-доминантный признак, является результатом развития закладок дополнительных пальцев, характерных в норме для далеких предковых форм. Известен феномен полифалангии, характеризующийся увеличением числа фаланг обычно большого пальца кисти. В основе его возникновения - развитие трех фаланг в первом пальце, как это в норме наблюдается у пресмыкающихся и земноводных с недифференцированными пальцами конечностей. Двусторонняя полифалангия наследуется аутосомно-доминантно.
Серьезным пороком развития является нарушение гетеротопии пояса верхних конечностей из шейной области на уровень 1-2-го грудных позвонков. Эту аномалию называют болезнью Шпренгеля или врожденным высоким стоянием лопатки (рис. 14.16). Она выражается в том, что плечевой пояс с одной либо с двух сторон находится выше нормального положения на несколько сантиметров. В связи с тем что такое нарушение часто сопровождается аномалиями рёбер, грудного отдела позвоночника и деформацией лопаток, следует думать, что механизмы его возникновения - не только нарушение перемещения органов, но и обусловленное этим нарушение морфогенетических корреляций (см. § 13.4).
Сравнительно-анатомический обзор эволюции скелета хордовых свидетельствует о том, что скелет человека полностью гомологичен опорному аппарату предковых и родственных форм. Поэтому многие пороки его развития у человека можно объяснить родством млекопитающих с пресмыкающимися, земноводными и рыбами. Однако в процессе антропогенеза появились такие особенности скелета, которые характерны лишь для человека и связаны с его прямохождением и трудовой деятельностью. К ним относят: 1) изменения стопы переставшей выполнять хватательную функцию, выражающиеся в потере способности к противопоставлению большого пальца и появлению ее сводов, служащих для амортизации при ходьбе; 2) изменения позвоночного столба - его S-образный изгиб, обеспечивающий пластичность движений в вертикальном положении; 3) изменения черепа - резкое уменьшение его лицевой части и увеличение мозговой, смещение большого затылочного отверстия кпереди, увеличение сосцевидного отростка и сглаживание затылочного рельефа, к которому прикрепляются мышцы шеи и выйная связка; 4) специализация верхних (передних) конечностей как органа труда; 5) появление подбородочного выступа в связи с развитием членораздельной речи.

Рис. 14.15. Формирование передних конечностей, их гетеротопия и иннервация в онтогенезе человека. А -врастание шейных миотомов в формирующуюся переднею конечность зародыша; Б -развитие кожной иннервации руки; В - расположение шейного и плечевого сплетений, участвующих в иннервации руки:
1 -шейные миотомы, 2- грудные миотомы, 3 -поясничные миотомы; буквами С, Т, L обозначены шейные, грудные и поясничные сегменты

Рис. 14.16. Болезнь Шпренгеля (пояснение см. в тексте)
Несмотря на то что становление анатомо-морфологических особенностей скелета у человека, по-видимому, завершено, адаптации к прямохождению у него имеют, как и все адаптации вообще, относительный характер. Так, при большой физической нагрузке возможно смещение позвонков или межпозвонковых дисков. Человек, перейдя к прямохождению, утратил способность к быстрому бегу и передвигается значительно медленнее большинства четвероногих животных.
Естественно, что в ходе внутриутробного развития черты скелета, характеризующие человека как уникальный биологический вид, формируются на конечных его этапах либо даже, как, например, S-образная форма позвоночника, в раннем постнатальном периоде развития. Они являются фактически анаболиями, возникшими в ходе филогенеза приматов. Поэтому атавистические аномалии скелета, связанные с задержками развития признаков, характерных только для человека, встречаются наиболее часто. Они практически не снижают жизнеспособность, но дети, обладающие ими, нуждаются в ортопедической коррекции, гимнастике и массаже. К таким аномалиям относят легкие формы врожденного плоскостопия, косолапости, узкую грудную клетку, отсутствие подбородочного выступа и некоторые другие.
Мышечная система
У представителей типа Хордовые мускулатура подразделяется по характеру развития и иннервации на соматическую и висцеральную.
Соматическая мускулатура развивается из миотомов и иннервируется нервами, волокна которых выходят из спинного мозга в составе брюшных корешков спинномозговых нервов. Висцеральная мускулатура развивается из других участков мезодермы и иннервируется нервами вегетативной нервной системы. Вся соматическая мускулатура поперечно-полосата, а висцеральная может быть как поперечно-полосатой, так и гладкой (рис. 14.17).

Рис. 14.17. Соматическая и висцеральная мускулатура позвоночных:
1 -соматическая мускулатура, развивающаяся из миотомов, 2- висцеральная мускулатура жаберной области
Висцеральная мускулатура
Наиболее существенные изменения претерпела висцеральная мускулатура, связанная с висцеральными дугами передней части пищеварительной трубки. У низших позвоночных большая часть этой мускулатуры представлена общим сжимателем висцерального аппарата - m. constrictor superficialis, покрывающим всю область жаберных дуг со всех сторон. В области челюстной дуги эта мышца иннервируется тройничным нервом (V), в области подъязычной дуги - лицевым (VII), в области первой жаберной дуги - языкоглоточным (IX), наконец, ее часть, лежащая каудальнее,- блуждающим нервом (X). В связи с этим все производные соответствующих висцеральных дуг и мышц, связанных с ними, иннервируются впоследствии у всех позвоночных перечисленными нервами.
В передней части сжимателя обособляется крупная мускулатурная масса, обслуживающая челюстной аппарат. Позади висцерального аппарата дифференцируется трапециевидная мышца m. trapezius, отдельными пучками прикрепляющаяся к последним жаберным щелям и переднему краю спинного отдела плечевого пояса. Часть поверхностного сжимателя в области подъязычной дуги у пресмыкающихся разрастается, охватывает шею снизу и с боков и образует сжиматель шеи m. sphincter colli. У млекопитающих эта мышца делится на два слоя: глубокий и поверхностный. Глубокий сохраняет прежнее название, а поверхностный называется platysma myoides и располагается подкожно. Эти две мышцы разрастаются на всю область головы и дают начало сложной системе лицевой подкожной мускулатуры, которую у приматов и человека называют мимической. Поэтому вся мимическ
ЛЕКЦИЯ МИОЛОГИЯ ФИЛОГЕНЕЗ, ОНТОГЕНЕЗ И ФУНКЦИОНАЛЬНАЯ АНАТОМИЯ МЫШЕЧНОЙ СИСТЕМЫ Выполнили: Владимирова Я. Б. Кокорева Т. В.

 Мышцы или мускулы (от лат. musculus - мышка, маленькая мышь) - органы тела животных и человека, состоящие из упругой, эластичной мышечной ткани, способной сокращаться под влиянием нервных импульсов. Предназначены для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 86, 3 % из воды. В теле человека 640 мышц
Мышцы или мускулы (от лат. musculus - мышка, маленькая мышь) - органы тела животных и человека, состоящие из упругой, эластичной мышечной ткани, способной сокращаться под влиянием нервных импульсов. Предназначены для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 86, 3 % из воды. В теле человека 640 мышц
 Мотивация: - - - возможности совершаемого движения, объём движения; активные или пассивные движения запускаются в действие той или иной мышечной группой; воздействуя на мышечный аппарат, мы изменяем общее состояние; мышечный рельеф является ориентиром для топографии сосудов и нервов; пересадка мышц, то есть мышца может «переучиваться» .
Мотивация: - - - возможности совершаемого движения, объём движения; активные или пассивные движения запускаются в действие той или иной мышечной группой; воздействуя на мышечный аппарат, мы изменяем общее состояние; мышечный рельеф является ориентиром для топографии сосудов и нервов; пересадка мышц, то есть мышца может «переучиваться» .
 Развитие мышц Краниального происхождения - из головных миотомов (склеротомов) и мезенхимы жаберных дуг. Иннервируются ветвями черепномозговых нервов Спинального происхождения - из миотомов туловищного отдела зародыша: из вентральных миотомов иннервируются передними ветвями СМН; - из дорзальных миотомов иннервируются задними ветвями СМН - Аутохтонные мышцы – мыщцы, оставшиеся на месте своей первичной закладки. Трункофугальные мышцы – мыщцы, переместившиеся с туловища на конечности. Трункопетальные мышцы – мыщцы, переместившиеся с конечностей на туловище.
Развитие мышц Краниального происхождения - из головных миотомов (склеротомов) и мезенхимы жаберных дуг. Иннервируются ветвями черепномозговых нервов Спинального происхождения - из миотомов туловищного отдела зародыша: из вентральных миотомов иннервируются передними ветвями СМН; - из дорзальных миотомов иннервируются задними ветвями СМН - Аутохтонные мышцы – мыщцы, оставшиеся на месте своей первичной закладки. Трункофугальные мышцы – мыщцы, переместившиеся с туловища на конечности. Трункопетальные мышцы – мыщцы, переместившиеся с конечностей на туловище.



 Поперечно-полосатая Гладкая 1. Единицей организации является миоцит. Длина около 50 мкм. Ширина от 6 мкм. 2. Непроизвольное сокращение Контроль со стороны автономной нервной системы Движение волнообразное, работает медленно, так как нервное волокно подходит НЕ к каждой клетке Медленно вступают в действие, но сохраняются на длительном протяжении Не имеет точной пространственной ориентации клеток 3. 4. 5. 6. 1. 2. 3. 4. 5. 6. Сердечная Единицей организации является мышечное волокно- множество миобластов, плавающих в общей цитоплазме (саркоплазме). У них общая сарколемма. Длина около 40 -100 мм. Ширина от 7 мм. Произвольное сокращение Контроль со стороны соматической нервной системы Быстрое сокращение, быстрая реакция, так каждое мышечное волокно имеет нервно -мышечный синапс Быстро включаются, но имеет кратковременное действие Четкая ориентация мышечных волокон
Поперечно-полосатая Гладкая 1. Единицей организации является миоцит. Длина около 50 мкм. Ширина от 6 мкм. 2. Непроизвольное сокращение Контроль со стороны автономной нервной системы Движение волнообразное, работает медленно, так как нервное волокно подходит НЕ к каждой клетке Медленно вступают в действие, но сохраняются на длительном протяжении Не имеет точной пространственной ориентации клеток 3. 4. 5. 6. 1. 2. 3. 4. 5. 6. Сердечная Единицей организации является мышечное волокно- множество миобластов, плавающих в общей цитоплазме (саркоплазме). У них общая сарколемма. Длина около 40 -100 мм. Ширина от 7 мм. Произвольное сокращение Контроль со стороны соматической нервной системы Быстрое сокращение, быстрая реакция, так каждое мышечное волокно имеет нервно -мышечный синапс Быстро включаются, но имеет кратковременное действие Четкая ориентация мышечных волокон

 Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани- эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.
Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани- эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.
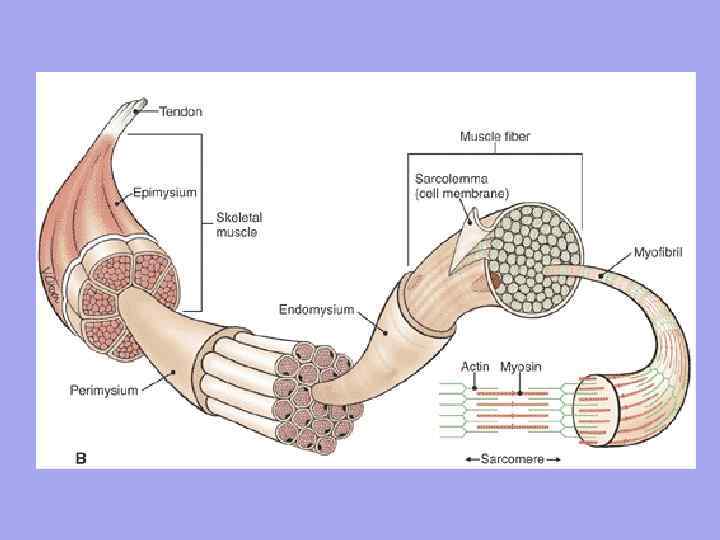
 В мышце как органе есть соединительная ткань Эндомизий – тонкая соединительная ткань, окружает каждое мышечное волокно и небольшие группы волокон. Перимизий – покрывает более крупные комплексы мышечных волокон и мышечные пучки.
В мышце как органе есть соединительная ткань Эндомизий – тонкая соединительная ткань, окружает каждое мышечное волокно и небольшие группы волокон. Перимизий – покрывает более крупные комплексы мышечных волокон и мышечные пучки.
 Значение эндомизия и перимизия 1. Через эндомизиум и перимизиум к мышечному волокну подходят сосуды и нервы. Образуют строму органа; 2. Мышечные волокна формируются в пучки, пучки в мышцу; 3. Так как эндомизий сращен с сарколеммой мышечного волокна, следовательно, сокращающее мышечное волокно может растягиваться лишь до определенного предела
Значение эндомизия и перимизия 1. Через эндомизиум и перимизиум к мышечному волокну подходят сосуды и нервы. Образуют строму органа; 2. Мышечные волокна формируются в пучки, пучки в мышцу; 3. Так как эндомизий сращен с сарколеммой мышечного волокна, следовательно, сокращающее мышечное волокно может растягиваться лишь до определенного предела
 Миофибриллы в волокне окружены оболочкой – сарколеммой, и погружены в специальную среду – саркоплазму. В зависимости от содержания пигмента и кислорода волокна делятся на белые и красные. Белые волокна –анаэробы, содержат больше миофибрилл, меньше саркоплазмы. Запускаются быстро, но долго работать не могут. Пример: грудинно-ключичнососцевидная, икроножная мышцы. Красные волокна – волокна толстые. Много миоглобина в саркоплазме и цитохрома в митохондриях, но меньше миофибрилл. Медленно запускаются, но работают долго. Пример: мышцы спины, диафрагма.
Миофибриллы в волокне окружены оболочкой – сарколеммой, и погружены в специальную среду – саркоплазму. В зависимости от содержания пигмента и кислорода волокна делятся на белые и красные. Белые волокна –анаэробы, содержат больше миофибрилл, меньше саркоплазмы. Запускаются быстро, но долго работать не могут. Пример: грудинно-ключичнососцевидная, икроножная мышцы. Красные волокна – волокна толстые. Много миоглобина в саркоплазме и цитохрома в митохондриях, но меньше миофибрилл. Медленно запускаются, но работают долго. Пример: мышцы спины, диафрагма.
 Каждая мышца имеет сеть кровеносных сосудов. Сокращения мышцы способствует току крови. В расслабленной неработающей мышце большая часть кровеносных капилляров закрыта для тока крови. При сокращении мышцы все кровеносные капилляры сразу открываются.
Каждая мышца имеет сеть кровеносных сосудов. Сокращения мышцы способствует току крови. В расслабленной неработающей мышце большая часть кровеносных капилляров закрыта для тока крови. При сокращении мышцы все кровеносные капилляры сразу открываются.

 Строение мышцы Каждая мышца одним концом соединяется с одной костью (начало мышцы), а другим – с другой (прикрепление мышцы). В мышце различают: головку брюшко хвостовую часть.
Строение мышцы Каждая мышца одним концом соединяется с одной костью (начало мышцы), а другим – с другой (прикрепление мышцы). В мышце различают: головку брюшко хвостовую часть.
 К каждому мышечному волокну подходят двигательные нервные волокна и отходят чувствительные нервные волокна Количество нервных окончаний в мышце зависит от степени функциональной активности мышц.
К каждому мышечному волокну подходят двигательные нервные волокна и отходят чувствительные нервные волокна Количество нервных окончаний в мышце зависит от степени функциональной активности мышц.
 Каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется моторной единицей. Характерно, что мышечные волокна, принадлежащие к одной моторной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.
Каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется моторной единицей. Характерно, что мышечные волокна, принадлежащие к одной моторной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.


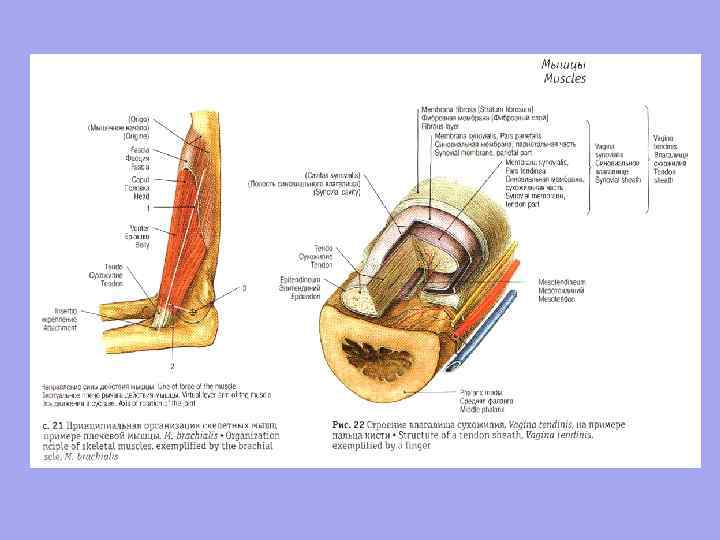
 Сухожилие – это плотный волокнистый соединительно-тканный тяж, которым мышца начинается или прикрепляется к скелету
Сухожилие – это плотный волокнистый соединительно-тканный тяж, которым мышца начинается или прикрепляется к скелету
 перитенониум коллагеновые волокна IV типа эндотенониум Коллагеновые волокна сухожилия, переплетающиеся с коллагеновыми волокнами надкостницы, вплетаются в основное вещество костной ткани, образуя на костях гребни, бугорки, бугристости, впадины, вдавления.
перитенониум коллагеновые волокна IV типа эндотенониум Коллагеновые волокна сухожилия, переплетающиеся с коллагеновыми волокнами надкостницы, вплетаются в основное вещество костной ткани, образуя на костях гребни, бугорки, бугристости, впадины, вдавления.
 Фасции – это соединительнотканные коллагенные волокна с небольшой примесью эластических волокон Поверхностная височная фасция Глубокие фасции бедра
Фасции – это соединительнотканные коллагенные волокна с небольшой примесью эластических волокон Поверхностная височная фасция Глубокие фасции бедра
 1. 2. 3. 4. 5. Фасции отделяют мышцы от кожи и устраняют смещение кожи при движениях сокращающихся мышц. Фасции экономят силу сокращения мышц, устраняя трение между мышцами во время сокращения. Фасции растягивают крупные вены при натяжении, в результате чего кровь с периферии «присасывается» в эти вены. Фасции имеют значение как барьеры, препятствующие распространению инфекции и опухолей. Во время операций фасции помогают определять расположение мышц, кровеносных сосудов, внутренностей.
1. 2. 3. 4. 5. Фасции отделяют мышцы от кожи и устраняют смещение кожи при движениях сокращающихся мышц. Фасции экономят силу сокращения мышц, устраняя трение между мышцами во время сокращения. Фасции растягивают крупные вены при натяжении, в результате чего кровь с периферии «присасывается» в эти вены. Фасции имеют значение как барьеры, препятствующие распространению инфекции и опухолей. Во время операций фасции помогают определять расположение мышц, кровеносных сосудов, внутренностей.
 Классификация мышц Скелетные мышцы разнообразны по форме, структуре, положению относительно осей суставов и т. д. , поэтому классифицируются по-разному.
Классификация мышц Скелетные мышцы разнообразны по форме, структуре, положению относительно осей суставов и т. д. , поэтому классифицируются по-разному.

 III. По функциональной особенности Статические (сильные) – короткое брюшко и длинное сухожилие. Мышцы работают с большей силой, но с меньшим размахом движения. Динамические (ловкие) – длинные мышечные пучки, сухожилия короткие. Мышцы работают с меньшей силой, но производят движения большего размаха
III. По функциональной особенности Статические (сильные) – короткое брюшко и длинное сухожилие. Мышцы работают с большей силой, но с меньшим размахом движения. Динамические (ловкие) – длинные мышечные пучки, сухожилия короткие. Мышцы работают с меньшей силой, но производят движения большего размаха
 Вспомогательный аппарат мышц Скелетные мышцы имеют вспомогательный аппарат, облегчающий их функционирование. n n n Фасции; Костно-фасциальные влагалища; Синовиальные сумки; Синовиальные влагалища сухожилий; Мышечные блоки; Сесамовидные кости.
Вспомогательный аппарат мышц Скелетные мышцы имеют вспомогательный аппарат, облегчающий их функционирование. n n n Фасции; Костно-фасциальные влагалища; Синовиальные сумки; Синовиальные влагалища сухожилий; Мышечные блоки; Сесамовидные кости.
 Аномалии развития мышц Встречаются очень часто и делятся на три группы: 1. Отсутствие какой-либо мышцы; 2. Наличие дополнительной мышцы, которой нет в природе. 3. Дополнительные пучки имеющейся мышцы.
Аномалии развития мышц Встречаются очень часто и делятся на три группы: 1. Отсутствие какой-либо мышцы; 2. Наличие дополнительной мышцы, которой нет в природе. 3. Дополнительные пучки имеющейся мышцы.
 Пороки развития Недоразвитие грудинно-ключично-сосцевидной мышцы – Кривошея Недоразвитие диафрагмы. Причина появление диафрагмальных грыж. Недоразвитие дельтовидной и трапециевидной мышц – Деформация плечевого пояса и плеча
Пороки развития Недоразвитие грудинно-ключично-сосцевидной мышцы – Кривошея Недоразвитие диафрагмы. Причина появление диафрагмальных грыж. Недоразвитие дельтовидной и трапециевидной мышц – Деформация плечевого пояса и плеча


 I. По форме: Веретенообразные; Лентовидные; Плоские широкие; Зубчатые; Длинные; n n n Квадратные; Треуголные; Круглые; Дельтовидная; Камбаловидная и т. д.
I. По форме: Веретенообразные; Лентовидные; Плоские широкие; Зубчатые; Длинные; n n n Квадратные; Треуголные; Круглые; Дельтовидная; Камбаловидная и т. д.



 II. По направлению мышечных волокон С прямыми параллельными волокнами; С поперечными; С круговыми; Перистые: A. Одноперистые; Двуперистые; C. Многоперистые. B.
II. По направлению мышечных волокон С прямыми параллельными волокнами; С поперечными; С круговыми; Перистые: A. Одноперистые; Двуперистые; C. Многоперистые. B.

 IV. По функции: Приводящие; Отводящие; Сгибающие; Разгибающие; Пронаторы; n n Супинаторы; Напрягающие; Мышцы – синергисты; Мышцы – антагонисты.
IV. По функции: Приводящие; Отводящие; Сгибающие; Разгибающие; Пронаторы; n n Супинаторы; Напрягающие; Мышцы – синергисты; Мышцы – антагонисты.
 V. По отношению к суставу: Односуставные; Двусуставные; Многосуставные.
V. По отношению к суставу: Односуставные; Двусуставные; Многосуставные.
Мышцы развиваются из мезодермы сомитов
склеротом, дерматом и миотом
Из дорсальных частей миотомов возникают глубокие, собственные вентральных аутохтонными (местными, туземными
трункофугальные
трункопетальные
· Филогенез мышечной системы
Развитие групп мышц
Развитие диафрагмы
Онтогенез мышечной системы: источники и сроки развития
Скелетные мышцы развиваются из мезодермы. У человеческого эмбриона примерно на 20-й день развития по бокам от нервного желобка возникают сомиты. Несколько позже в сомитах можно выделить их часть - миотомы. Клетки миотомов становятся веретенообразными и превращаются в делящиеся миобласты. Часть миобластов дифференцируется. Другая часть миобластов остается недифференцированной и
превращается в миосателлитоциты. Некоторые миобласты контактируют между собой своими полюсами, затем в зонах контактов плазмалеммы разрушаются, а клетки объединяются между собой, формируя симпласты. К ним мигрируют недифференцированные миобласты, которые окружаются единой с миосимпластом базальной мембраной. Если мышцы туловища развиваются из дорсального отдела мезодермы (сегментированного), то висцеральные, мимические, жевательные и некоторые мышцы шеи, а также промежности - из несегментированного вентрального отдела мезодермы, располагающегося соответственно в головном или хвостовом концах тела (табл. 33). Из мезодермы зачатков конечностей образуются их аутохтонные (туземные) мышцы (греч. autos . сам, chton - земля). Ряд мышц также закладывается в зачатках конечностей, но впоследствии их проксимальные концы прикрепляются к костям туловища - это трункопетальные (лат. truncus - туловище, petere -направляться), например большая и малая грудные мышцы. В отличие от них, трункофугальные (лат. fugere - бегать) развиваются из миотомов туловища, но их дистальные концы прикрепляются к костям конечностей, например большая и малая ромбовидные мышцы.
Развитие из мезодермы
Деление на сомиты
Производные миотома: из дорзального участка развиваются мышцы спины
Из вентрального - мышцы груди и живота
Мезенхима - мышцы конечностей
I висцеральная дуга (ВД) - жевательные мышцы
II ВД - мимические мышцы
III и IV ВД - мышцы мягкого неба, глотки, гортани, верхней части пищевода
V ВД - грудино-ключично-сосцевидная и трапециевидная мышцы
Из затылочных миотомов - мышцы языка
Из предушных миотомов - мышцы глазного яблока
Мышца: определение, строение.
Мускул (мышца) – орган, построенный из мышечных волокон (клеток), каждое из них обладает соединительно-тканной оболочкой – эндомизием . В пучки мышечные волокна объединяет другая фиброзная оболочка – перимизий , а весь мускул заключается в общую фиброзную оболочку, образованную фасцией – эпимизий . Между пучками проходят сосуды и нервы, снабжающие мышечные волокна.
На макроуровне скелетная мышца имеет:
· брюшко (вентер) – мясистая часть органа, занимающая его средину;
· сухожилие (тендо), относящееся к дистальному концу, оно может быть в виде апоневроза, сухожильных перемычек, длинных пучков продольных фиброзных волокон;
· головку , составляющую проксимальную часть;
· сухожилие и головка крепятся на противоположных концах костей.
Мышечные волокна располагаются параллельными рядами и соединяются в пучки, которые образуют саму скелетную мышцу. Небольшие мышечные пучки покрыты тонкой соединительной тканью - эндомизием (endomysium), крупные - перимизием (perimysium), а всю мышцу в целом покрывает плотная соединительная ткань - эпимизий (epimysium). На концах мышцы переходят в сухожилия, которые обладают большей эластичностью, упругостью и прочностью, чем мышечные волокна, что позволяет избегать разрывов мышц и их отрывов от костей при интенсивной внутренней нагрузке или сильном внешнем воздействии.
Волокна составляют примерно 86-90% от общей массы мышцы. Остальные проценты делят между собой кровеносные сосуды и нервы, обеспечивающие трофику (жизнедеятельность), питание и работоспособность мышц.
В мышце выделяют головку (caput) - начальную часть, брюшко (venter) - среднюю часть и хвост (cauda) - конечную часть (рис. 91). От длины мышцы зависит степень размаха, который она может обеспечить. У каждой мышцы есть точка начала (origo) и место крепления (insertio).
Классификация мышц по происхождению, строению, форме и функции.
В теле человека насчитывается около 400 мышц. Они имеют разную форму, размеры, местоположение, функции. Классификация мышц возможна по разным принципам:
По фоме различают мышцы:
короткие
Длинные мышцы встречаются в основном на конечностях, имеют веретеновидную форму; начало таких мышц называется головка, а прикрепление (конец) - хвост. Сухожилия таких мышц имеют вид длинных лент. Некоторые длинные мышцы имеют несколько головок (две, три, четыре и называются соответственно двуглавыми, трехглавыми, четырехглавыми). Бывают мышцы не с одним, а с несколькими брюшками, которые
соединяются сухожилиями; они называются многобрюшными. Бывают многохвостые мышцы, например, сгибатели пальцев.
Широкие мышцы располагаются преимущественно на туловище. Короткие мышцы сходны по внешней форме либо с длинными, либо с широкими мышцами, но имеют небольшие размеры.
По направлению волокн различают мышцы:
с прямыми параллельными волокнами
с косыми волокнами
с круговыми волокнами (окружают отверстия).
По местоположению мышцы делятся на:
поверхностные и глубокие; наружные и внутренние
мышцы туловища
мышцы головы
мышцы шеи; мышцы конечностей.
По функциям мышцы бывают:
сгибатели - разгибатели
приводящие - отводящие
вращатели внутрь или наружу
замыкатели (сфинктеры) - расширители
подниматели-опускатели
Закономерности расположения мышц.
· Соответственно строению тела по принципу двусторонней симметрии мышцы являются парными или состоят из двух симметричных половин.
· Мышцы располагаются между точками прикрепления по кратчайшему расстоянию.
· Мышца перекидывается по крайней мере через один сустав (но может быть и больше) и как правило перпендикулярно оси, через которую происходит движение сустава.
Анатомический и физиологический поперечник мышц: определение понятий.
Анатомический поперечник характеризует величину мышцы: длину, ширину, толщину. Под ним понимается площадь поперечного сечения всей мышцы, проходящая в наиболее широкой части брюшка перпендикулярно к длинной оси.
Физиологический поперечник характеризует силу мышцы, поэтому под ним понимают суммарную площадь поперечного сечения всех мышечных волокон.
Работа мышц.
Динамическая работа (характеризуется изотоническим сокращением мышц):
Преодолевающая – работа, при которой сокращение мышцы ведет изменению положения части тела за счет преодоления тяжести данной части тела или какого-либо сопротивления (например, груза);
Уступающая – работа, при которой сила мышцы уступает действию силы тяжести тела или его части и удерживаемого груза;
Баллистическая работа характеризуется быстрым и резким сокращением предварительно растянутой мышцы с последующим движением звена по инерции.
Статическая работа (характеризуется изометрическим сокращением мышц):
Удерживающая – работа, при которой сокращение мышцы удерживает тело или груз в определенном положении без перемещения в пространстве.
Области и треугольники шеи
Области шеи:
I. Передняя область шеи - треугольники: 1. Поднижнечелюстной; 2. Сонный; 3.Лопаточно-трахеальный;
II. Боковая область шеи - треугольники: 4. Лопаточно-ключичный; 5. Лопаточно-трапециевидный.
III. Задняя область шеи.
Верхняя шейная граница проходит через подбородок, основание нижней челюсти и по заднему краю ее ветвей, через височно-нижнечелюстной сустав, вершину сосцевидного отростка и далее по верхней выйной линии.
Нижняя – через яремную вырезку грудины, грудино-ключичный сустав, ключицу, ключично-акромиальный сустав и сзади по линии акромиона - остистый отросток выступающего (YII) позвонка. Фронтальная плоскость, проведенная на уровне поперечных отростков и тел шейных позвонков, делит шею на переднюю и заднюю (выйную) области. Между ними выделяют латеральную и грудино-ключично-сосцевидную области. Передние мышцы шеи своеобразно перекрещиваются и образуют много треугольников.
Передняя область или передний треугольник шеи ограничен по бокам передними краями грудино-ключично-сосцевидных мышц, вверху – подбородком, основанием и ветвями нижней челюсти, сосцевидными отростками, внизу – яремной вырезкой грудины.
Передняя срединная линия от подбородка до яремной вырезки делит область на медиальные треугольники : правый и левый.
В каждом медиальном треугольнике различают вверху: поднижнечелюстной треугольник , ограниченный передним и задним брюшками двубрюшных мышц и нижней челюстью. В нем располагается поднижнечелюстная слюнная железа и маленький язычный треугольник, описанный Н. И. Пироговым в границах:
· передней – задний край челюстно-подъязычной мышцы,
· задней - нижний край заднего брюшка двубрюшной мышцы;
· верхней – подъязычный нерв;
· площадь треугольника занимает подъязычно-язычная мышца и лежащая под ней язычная артерия , для оперативного доступа к которой и был выделен Н.И. Пироговым данный треугольник.
Середину передней области составляет каротидный (сонный ) треугольник , образованный спереди и снизу верхним брюшком лопаточно-подъязычной мышцы, сверху – задним брюшком двубрюшной мышцы, а сзади – передним краем грудино-ключично-сосцевидной.
В сонном треугольнике проходят внутренняя яремная вена, блуждающий нерв и общая сонная артерия , которая в его пределах делится на уровне верхнего края щитовидного хряща на наружную и внутреннюю. В нижней части треугольника общая сонная артерия прилежит к переднему бугорку поперечного отростка YI шейного позвонка и к нему (сонный бугорок) ее прижимают при прощупывании пульса и остановке кровотечения.
Нижнюю часть передней области занимает лопаточно-трахеальный треугольник в границах: верхнелатеральной – верхнее брюшко лопаточно-подъязычной мышцы, задненижней – край грудино-ключично-сосцевидной мышцы, медиальной - передняя срединная линия. В глубине треугольника лежат трахея и пищевод.
Грудино-ключично-сосцевидная область соответствует одноименной мышце и служит хорошим ориентиром между латеральным и медиальным треугольником. Передний край мышцы соответствует проекционной линии каротидной артерии, яремной внутренней вены и блуждающего нерва, расположенного между ними.
Латеральная область шеи имеет переднюю границу по заднему краю грудино-ключично-сосцевидной мышцы, заднюю по трапециевидной мышце, нижнюю – по ключице.
В ней находятся.
· Лопаточно-трапециевидный треугольник, занимающий верхний отдел, располагается между краями трапециевидной, грудино-ключично-сосцевидной мышц (боковые стороны) и нижним брюшком лопаточно-подъязычной мышцы (нижняя сторона). В нем проецируется шейное сплетение и его короткие ветви.
· Лопаточно-ключичный треугольник образован ключицей (нижняя сторона) и краями грудино-ключично-сосцевидной, лопаточно-подъязычной (нижнее брюшко) мышц. Внутри его – в лестничных промежутках - находится горизонтальный сосудисто-нервный пучок шеи в составе (спереди и назад) подключичных вены, артерии и стволов плечевого сплетения.
Задняя область шеи имеет верхнюю границу по верхней выйной линии, боковые границы – по передним краям трапециевидной мышцы, нижнюю – по линии акромион-остистый отросток YII шейного позвонка. Область занята многослойной задней мышечной группой, описанной выше. Под затылком в задней области находится подзатылочный треугольник, ограниченный задними прямыми и косыми мышцами головы.
Слабые места диафрагмы
Диафрагма – это непарная куполообразной формы мышечно-сухожильная мембрана, разделяющая грудную и брюшную полость.
Слабые места диафрагмы:
1. Грудино-реберный треугольник
2. Пояснично-реберный треугольник
Паховый канал.
Паховый канал: это щелевидный промежуток, расположенный между глубоким и поверхностным паховыми кольцами.Стенки пахового канала: передняя стенка – апоневроз наружной косой мышцы живота, задняя – поперечная фасция, верхняя – нижние пучки внутренней косой мышцы живота и поперечной мышцы живота, нижняя стенка – паховая связка.
Отверстия пахового канала:
Поверхностное паховое кольцо. Границы: сверху – медиальная ножка, снизу – латеральная ножка, латерально – межножковые волокна, медиально – загнутая связка.
Глубокое паховое кольцо (внутреннее отверстие пахового канала) расположено на задней стенке пахового канала.
Приводящий канал.
Бедренно-подколенный (приводящий) канал образован следующими структурами:
· медиальная стенка - большая приводящая мышца;
· латеральная - медиальная широкая мышца;
· передняя - фиброзная пластинка (ламина васто-аддукториа) - из глубокого листка широкой фасции, натянутая между вышеназванными мышцами.
· Входное (верхнее) отверстие канала лежит под портняжной мышцей, выходное (нижнее) находится в подколенной ямке в виде щели сухожилия большого аддуктора; переднее отверстие располагается в фиброзной пластинке (вастоаддукторной) на уровне нижней трети бедра. Нижнее отверстие (выход из канала) открывается в подколенную ямку.
Через подвздошно-гребенчатую, бедренную борозды и приводящий канал проходят бедренные артерия, вена, большой скрытый нерв, причем скрытый нерв и ветвь бедренной артерии – нисходящая коленная, - покидают канал через переднее отверстие.
Каналы голени.
· Голенно-подколенный канал Грубера проходит от подколенной ямки до медиальной лодыжки. Переднюю стенку его образуют задняя большеберцовая мышца и длинный сгибатель большого пальца, заднюю – камбаловидная мышца. В канале проходят задняя большеберцовая артерия и вены, большеберцовый нерв. Через его переднее отверстие вверху межкостной перепонки выходит передняя большеберцовая артерия и сопровождающие вены.
· В средней трети голени от него ответвляется в латеральном направлении нижний мышечно-малоберцовый канал. Он находится между фибулой и длинным сгибателем большого пальца. В нем проходят малоберцовые артерия и вена.
· Верхний мышечно-малоберцовый канал находится между длинной малоберцовой мышцей и фибулой, содержит поверхностный малоберцовый нерв.
Филогенез мышечной системы: закономерности развития.
Мышцы развиваются из мезодермы . На туловище они возникают из первичной сегментированной мезодермы – сомитов : 3-5 затылочных, 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых, 4-5 копчиковых.
Каждый сомит подразделяется на склеротом, дерматом и миотом – из него и развиваются мышцы туловища.
Сомиты появляются рано, когда длина зародыша составляет 10-15 мм.
Из дорсальных частей миотомов возникают глубокие, собственные (аутохтонные) мышцы спины, из вентральных – глубокие мышцы груди и живота. Они закладываются, развиваются и остаются в пределах туловища – поэтому называются аутохтонными (местными, туземными ). Очень рано миотомы связываются с нервной системой и каждому мышечному сегменту соответствует нервный сегмент. Каждый нерв следует за развивающейся мышцей, врастает в нее и, пока она не дифференцировалась, подчиняет своему влиянию.
В процессе развития часть скелетных мышц перемещается с туловища и шеи на конечности – трункофугальные мышцы: трапециевидная, грудино-ключично-сосцевидная, ромбовидные, поднимающая лопатку и др.
Часть мышц, наоборот, направляется с конечностей на туловище – трункопетальные мышцы: широчайшая мышца спины, большая и малая грудные, большая поясничная.
· Филогенез мышечной системы
Не изолированная мышечная система
Единый кожно-мускульный мешок
Появление поперечно-полосатой мышечной ткани
Разделение мышечных тяжей на миотомы
Развитие групп мышц
Развитие мышц конечностей (изменение среды обитания)
Развитие диафрагмы
Развитие всех групп мышц - выполнение дифференцированных движений
